Team der Back Clinic Anti Aging Chiropraktik und Funktionelle Medizin. Unser Körper befindet sich in einem ständigen und nie endenden Kampf ums Überleben. Zellen werden geboren, Zellen werden zerstört. Wissenschaftler schätzen, dass jede Zelle über 10,000 einzelnen Angriffen durch reaktive Sauerstoffspezies (ROS) oder freie Radikale standhalten muss. Ohne Fehler verfügt der Körper über ein unglaubliches Selbstheilungssystem, das dem Angriff standhält und beschädigtes oder zerstörtes wieder aufbaut. Das ist das Schöne an unserem Design.
Die Biologie des Alterns verstehen und wissenschaftliche Erkenntnisse in Interventionen umsetzen, die die Gesundheit im späteren Leben durch Behandlungen verbessern. Es ist nützlich, eine klare, einheitliche Meinung darüber zu haben, was genau eine Anti-Aging-Behandlung ausmacht.
Schon vor den Tagen von Ponce de Leons Streben nach Langlebigkeit wurde der Mensch immer von der Chance auf ewige Jugend verführt. Die Chiropraktik mit ihrer Gesundheitsbewegung ist eine wirkungsvolle Methode zur Stabilisierung und Förderung dieser Selbstheilungskräfte. Dr. Alex Jimenez diskutiert Konzepte rund um die Anti-Aging-Pandora.
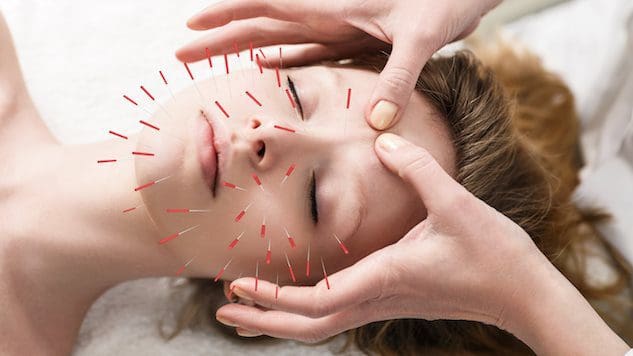
Kann die Einbeziehung von Akupunktur bei Personen, die die Hautgesundheit verbessern oder erhalten möchten, dazu beitragen, die Haut zu verbessern und den Alterungsprozess zu bekämpfen?
Kosmetische Akupunktur
Die kosmetische Akupunktur folgt der traditionellen Akupunkturpraxis des Einführens einer Nadel. Ziel ist es, Zeichen der Hautalterung umzukehren und die Hautgesundheit zu verbessern. Es wird manchmal als Akupunktur-Gesichtsverjüngung bezeichnet und wird als Alternative zu chirurgischen Facelifts und anderen herkömmlichen Verfahren eingesetzt. Vorläufige Studien haben untersucht, wie es dabei helfen kann, Altersflecken zu entfernen, schlaffe Augenlider zu straffen und Falten zu reduzieren. (Younghee Yun et al., 2013)
Wie Akupunktur funktioniert
In der Traditionellen Chinesischen Medizin oder TCM wird Akupunktur seit langem zur Verbesserung des Energieflusses – Qi oder Chi – im gesamten Körper eingesetzt. Es wird angenommen, dass diese Energie über Energiebahnen zirkuliert, die als Meridiane bekannt sind. Bei gesundheitlichen Problemen liegen laut TCM Störungen oder Blockaden im Kreislauf vor.
Akupunkteure können durch das Einführen von Nadeln in bestimmte Akupunkturpunkte eine optimale Durchblutung/Fluss wiederherstellen und die Gesundheit verbessern. (Nationale Gesundheitsinstitute, 2007)
Kosmetische Akupunktur
Kosmetische Akupunktur soll die Hautgesundheit verbessern und als Anti-Aging-Behandlung wirken, indem sie die Kollagenproduktion anregt. Dieses Protein ist ein Hauptbestandteil der Haut. Die innere Hautschicht verliert mit zunehmendem Alter an Kollagen und Festigkeit. Es bedarf jedoch weiterer Forschung, um die Behauptung zu untermauern, dass Akupunktur die Kollagenproduktion fördern kann. Einige schlagen vor, dass kosmetische Akupunktur zur Verjüngung der Haut beiträgt, indem sie die Gesamtenergie des Körpers verbessert. Eine Studie ergab, dass Einzelpersonen nach fünf Sitzungen kosmetischer Akupunktur im Gesicht Verbesserungen sahen. (Younghee Yun et al., 2013) Für optimale Ergebnisse wird jedoch empfohlen, ein- bis zweimal pro Woche zehn Behandlungen durchzuführen. Danach werden alle vier bis acht Wochen Erhaltungsbehandlungen durchgeführt. Im Gegensatz zu Botox oder Hautfüllern ist kosmetische Akupunktur keine schnelle Lösung. Der Fokus liegt auf der nachhaltigen Veränderung von Haut und Körper, d.h. einer Verbesserung von:
Wenn die Nadeln in die Haut eingeführt werden, entstehen Wunden, die als positive Mikrotraumata bekannt sind. Die natürlichen Heilungs- und Reparaturfähigkeiten des Körpers werden aktiviert, wenn er diese Wunden wahrnimmt. Diese Einstiche stimulieren das Lymph- und Kreislaufsystem, das die Hautzellen mit Nährstoffen und Sauerstoff versorgt und sie von innen heraus nährt.
Dies hilft, den Teint auszugleichen und die Ausstrahlung der Haut zu fördern.
Die positiven Mikrotraumata stimulieren auch die Produktion von Kollagen.
Dies trägt zur Verbesserung der Elastizität bei und minimiert Linien und Fältchen.
Alternativen
Mehrere natürliche Heilmittel können zur Verbesserung der Hautgesundheit beitragen und Anti-Aging-Vorteile bieten. Ceramide sind ein Fettmolekül, das natürlicherweise in der obersten Hautschicht vorkommt und ein Bestandteil von Hautpflegeprodukten ist. Diese können vor altersbedingter Trockenheit der Haut schützen. (L Di Marzio 2008) Vorläufige Untersuchungen deuten darauf hin, dass die Anwendung von weißem Tee auf die Haut den Abbau von Kollagen und Elastin bekämpfen kann – ein Protein, das die Elastizität der Haut unterstützt und ein Erschlaffen verhindert. Es gibt auch Hinweise darauf, dass natürliche Substanzen wie Arganöl, Borretschöl und Sanddorn feuchtigkeitsspendende Eigenschaften haben, die die Haut verbessern könnten.(Tamsyn SA Thring et al., 2009)
Während weitere Beweise für kosmetische Akupunktur erforderlich sind, kann die Integration von Akupunktur dabei helfen, Stress zu bewältigen und die allgemeine Gesundheit zu verbessern. Personen, die eine kosmetische Akupunktur in Betracht ziehen, sollten ihren Hausarzt konsultieren, um herauszufinden, ob diese für sie geeignet ist.
Gemeinsam die Gesundheit verbessern: Multidisziplinäre Bewertung und Behandlung
Bibliographie
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS und Choi, I. (2013). Wirkung kosmetischer Gesichtsakupunktur auf die Gesichtselastizität: eine offene, einarmige Pilotstudie. Evidenzbasierte Komplementär- und Alternativmedizin: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Das Nationale Zentrum für Komplementär- und Alternativmedizin. (2007). Akupunktur: Eine Einführung. Website des Nationalen Zentrums für Komplementär- und Alternativmedizin. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021). Zuverlässigkeit und Gültigkeit des Gesichtskontrollblatts (FCS): Checkliste für Selbstzufriedenheit mit kosmetischer Akupunktur. Arzneimittel (Basel, Schweiz), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG und Giuliani, M. (2008). Anstieg des Haut-Ceramid-Spiegels bei älteren Probanden nach einer kurzfristigen topischen Anwendung der bakteriellen Sphingomyelinase aus Streptococcus thermophilus. Internationale Zeitschrift für Immunpathologie und Pharmakologie, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P. & Naughton, DP (2009). Anti-Kollagenase-, Anti-Elastase- und antioxidative Wirkung von Extrakten aus 21 Pflanzen. BMC Komplementär- und Alternativmedizin, 9, 27. doi.org/10.1186/1472-6882-9-27
Wenn die Wirbelsäule eines Menschen in Topform bleibt, bedeutet dies weniger Schmerzen und mehr Beweglichkeit, Flexibilität und Freiheit. Der Verschleiß des Körpers ist ein natürlicher Effekt des Alterns, der jedem einzelnen von uns widerfährt. Wirbelsäulenprobleme im Zusammenhang mit dem Alter können schwerwiegend werden, wenn sie nicht durch Übungen, Dehnübungen und chiropraktische Maßnahmen angegangen und behandelt werden.
Altern und der Rücken
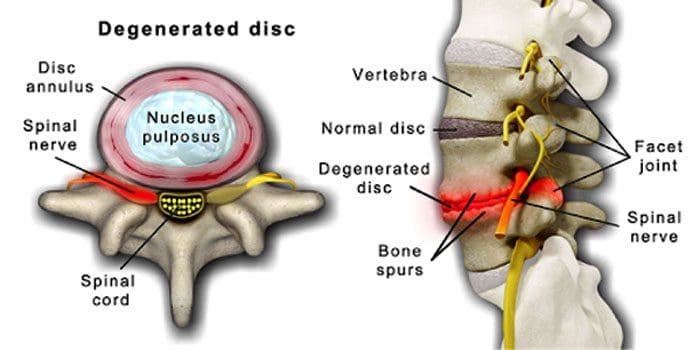
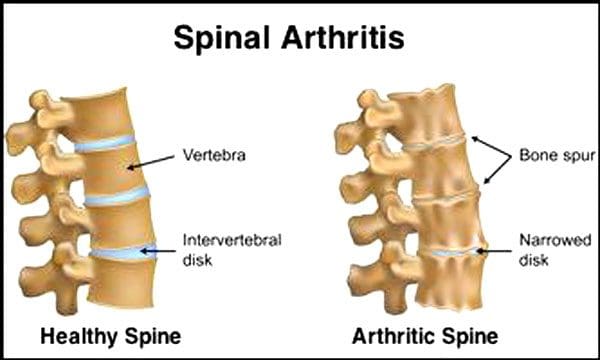
Es ist normal, dass sich die Bandscheiben und Gelenke mit zunehmendem Alter verschlechtern. Auch eine Spinalkanalstenose oder eine Verengung des Wirbelkanals kann Teil des Alterungsprozesses sein. Zwei durch Alterung hervorgerufene Zustände sind degenerativen Bandscheibenerkrankungen und Arthritis dazu kann auch gehören Versteifung der Wirbelsäulenbänder und Osteoporose.
Eine degenerative Bandscheibenerkrankung tritt bei 40 % der Menschen im Alter von 40 Jahren auf
Erhöht sich auf 80 % für Personen ab 80 Jahren.
Es dreht sich um Bandscheiben, die sich allmählich von überwiegend Wasser zu überwiegend Fett verändern.
Bei Fett verengen sich die Bandscheiben und verlieren an Elastizität.
Das sagen die Centers for Disease Control and Prevention 23 % der amerikanischen Erwachsenen leiden an Arthritis. Hierbei handelt es sich um eine Erkrankung, die hauptsächlich die Facettengelenke betrifft. Die Gelenke schwellen an, was den Bewegungsumfang einschränkt und die Spinalnerven beeinträchtigen kann, was zu Schmerzen, Schwäche und Ischiasbeschwerden führt. Mit der Zeit versteifen sich die Bänder um und in der Wirbelsäule, wodurch der Bewegungsbereich eingeschränkt wird und es zu einer Stenose kommt. Knochenschwund oder Osteoporose wird durch Veränderungen der Hormone und anderer Faktoren wie der Ernährung verursacht. Altern ist ein natürlicher Prozess, aber jeder Einzelne kann dazu beitragen, dass seine Wirbelsäule in Topform bleibt, egal wie alt er ist.
Eine gesunde Körperhaltung üben
Sofort richtige gesunde Körpermechanik ist ein Muss. Wenn Sie sich der Körperhaltung bewusst und achtsam sind, bleibt die Ausrichtung erhalten und der Körper bleibt im Gleichgewicht. Eine gesunde Haltung trägt dazu bei, die Auswirkungen von Folgendem zu reduzieren:
Spinalstenose
Degenerative Bandscheibenerkrankungen
Herniation
Gefahr von Wirbelsäulenfrakturen
Zum Üben der richtigen Körperhaltung gehört:
Reduzieren Sie das Herumhängen
Stellen Sie sicher, dass der Arbeitsplatz in Topform und ergonomisch einwandfrei ist
Dieser Ansatz lässt sich auch auf das Heben übertragen.
Achten Sie beim Heben darauf, die Knie zu beugen und die Wirbelsäule möglichst aufrecht zu halten.
Yoga
Yoga kann für eine gesündere, jugendlichere Wirbelsäule von großem Nutzen sein. Yoga erfüllt drei Bereiche, um die Wirbelsäule in Topform zu halten. Das beinhaltet:
Regelmäßiges Training
Bewahrt die Flexibilität
Erreicht das ideale Körpergewicht
Yoga ist eine Anti-Aging-Aktivität für die Wirbelsäule. Weil es:
Behält die Stärke bei
Flexibilität
Körperhaltung
Balance
Kann bei einer Vielzahl von Wirbelsäulenbeschwerden hilfreich sein, insbesondere bei Arthritisschmerzen
Stürze können schwere Verletzungen verursachen. Yoga kann auch dabei helfen, das Gleichgewicht zu verbessern.
Suchen Sie einen Chiropraktiker auf
Präventive Medizin ist der Schlüssel dazu, den Körper gesund, jugendlich und so stark wie möglich zu halten. Eine chiropraktische Untersuchung kann feststellen, ob Wirbelsäulenprobleme vorliegen und eine Diagnose stellen, um einen optimalen Behandlungsplan zu entwickeln. Wenn die Körperfunktionen aufgrund von Schmerzen im Rücken und/oder in den Beinen eingeschränkt sind, Kontaktieren Sie die Injury Medical Chiropractic and Functional Medicine Clinic und bringen Sie Ihre Wirbelsäule wieder in Topform.
Körperzusammensetzung
Übungs-/Stabilitätsballcurls
Diese Übung Trainiert Muskelgruppen, die für die Stärke der Wirbelsäule spezifisch sind, und umfasst Folgendes:
Beinbeuger
Gesäß
Tiefe Bauchmuskeln
Hüftabduktoren und -rotatoren
Übungen wie diese sind eine der effektivsten Methoden, um funktionelle Kraft und Ausdauer in den Oberschenkelmuskeln und Hüften aufzubauen und Verletzungen vorzubeugen. Um dieses Training durchzuführen:
Legen Sie sich mit gebeugten Knien auf den Rücken
Heben Sie die Beine an, sodass die Unterseite der Füße auf einem Gymnastikball ruht
Rollen Sie Ihre Beine aus, bis sie gerade sind
Halten Sie die Position ein oder zwei Sekunden lang
Kehren Sie zum oberen Ende der Bewegung zurück und spannen Sie gleichzeitig die Oberschenkelmuskulatur an
Wenn Sie diese Muskeln trainieren, werden Hock-, Ausfall- und Beugebewegungen für die Wirbelsäule einfacher.
Haftungsausschluss für den Blog-Beitrag von Dr. Alex Jimenez
Der Umfang unserer Informationen beschränkt sich auf Artikel, Themen und Diskussionen in den Bereichen Chiropraktik, Bewegungsapparat, physikalische Medizin, Wellness und sensible Gesundheit sowie auf funktionelle Medizin. Wir verwenden funktionelle Gesundheits- und Wellnessprotokolle zur Behandlung und Unterstützung der Behandlung von Verletzungen oder Störungen des Bewegungsapparates. Unsere Beiträge, Themen, Themen und Erkenntnisse decken klinische Fragen, Probleme und Themen ab, die sich direkt oder indirekt auf unseren klinischen Anwendungsbereich beziehen und diesen unterstützen. *
Unser Büro hat einen vernünftigen Versuch unternommen, unterstützende Zitate bereitzustellen, und die relevante Forschungsstudie oder Studien identifiziert, die unsere Stellen unterstützen. Auf Anfrage stellen wir dem Vorstand und / oder der Öffentlichkeit auch Kopien unterstützender Forschungsstudien zur Verfügung. Wir verstehen, dass wir Angelegenheiten abdecken, die eine zusätzliche Erklärung erfordern, wie sie in einem bestimmten Pflegeplan oder Behandlungsprotokoll hilfreich sein können. Um das oben genannte Thema weiter zu diskutieren, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900. Die in Texas und New Mexico lizenzierten Anbieter *
Die Lebensmittel, die wir essen, können unserer Gesundheit zuträglich oder schädlich sein. Eine schlechte Ernährung kann eine Vielzahl von Gesundheitsproblemen verursachen, darunter Fettleibigkeit, Herz-Kreislauf-Erkrankungen und Typ-2-Diabetes. Unterdessen kann die richtige Ernährung dazu führen, dass Sie sich energiegeladen fühlen, das Risiko von Gesundheitsproblemen verringern und dabei helfen, ein gesundes Gewicht zu halten und zu regulieren. Wenn Sie ein langes Leben fördern möchten, müssen Sie Ihren Körper mit guten Lebensmitteln versorgen. Im folgenden Artikel werden wir einige gute Lebensmittel auflisten, die letztendlich dazu beitragen können, die Langlebigkeit zu fördern, indem sie auch zur Verbesserung der allgemeinen Gesundheit und des Wohlbefindens beitragen.
Kreuzblütler
Kreuzblütler haben die einzigartige Fähigkeit, unsere Hormone zu verändern, das natürliche Entgiftungssystem des Körpers anzuregen und sogar das Wachstum von Krebszellen zu reduzieren. Diese müssen gründlich gekaut oder zerkleinert, gehackt, entsaftet oder gemixt gegessen werden, um ihre wohltuenden Eigenschaften freizusetzen. Es wurde auch festgestellt, dass Sulforaphan, das in Kreuzblütlern vorkommt, dabei hilft, die Blutgefäßwand vor Entzündungen zu schützen, die Herzerkrankungen verursachen können. Kreuzblütler wie Grünkohl, Kohl, Rosenkohl, Blumenkohl und Brokkoli gehören zu den nährstoffreichsten Lebensmitteln der Welt.
Salatgemüse
Rohes Blattgemüse hat weniger als 100 Kalorien pro Pfund, was es zum perfekten Lebensmittel zum Abnehmen macht. Der Verzehr von mehr Salat wird auch mit einem geringeren Risiko für Herzinfarkt, Schlaganfall, Diabetes und verschiedene Krebsarten in Verbindung gebracht. Rohes Blattgemüse ist außerdem reich an dem essentiellen B-Vitamin Folat sowie Lutein und Zeaxanthin, Carotinoiden, die zum Schutz der Augen beitragen können. Fettlösliche sekundäre Pflanzenstoffe wie Carotinoide, die in Salaten wie Kopfsalat, Spinat, Grünkohl, Grünkohl und Senfgrün enthalten sind, wirken ebenfalls antioxidativ und entzündungshemmend auf den Körper.
Nüsse
Nüsse sind ein Lebensmittel mit niedrigem glykämischen Index und eine großartige Quelle für gesunde Fette, Pflanzenproteine, Ballaststoffe, Antioxidantien, Phytosterine und Mineralien. Sie tragen auch dazu bei, die glykämische Last einer gesamten Mahlzeit zu reduzieren, was sie zu einem wesentlichen Bestandteil eines Anti-Diabetes-Mittels macht Diät. Unabhängig von ihrer Kaloriendichte kann der Verzehr von Nüssen zur Gewichtsabnahme beitragen. Nüsse können auch den Cholesterinspiegel senken und das Risiko von Herzerkrankungen verringern.
Saat
Samen liefern, ähnlich wie Nüsse, auch gesunde Fette, Antioxidantien und Mineralien, diese enthalten jedoch mehr Protein und sind reich an Spurenelementen. Chia-, Lein- und Hanfsamen sind reich an Omega-3-Fetten. Chia-, Lein- und Sesamsamen sind ebenfalls reich an Lignanen oder brustkrebsbekämpfenden Phytoöstrogenen. Darüber hinaus sind Sesamsamen reich an Kalzium und Vitamin E und Kürbiskerne reich an Zink.
Beeren
Beeren sind antioxidantienreiche Früchte, die zur Förderung der Herzgesundheit beitragen können. Forschungsstudien, bei denen Teilnehmer mehrere Wochen lang täglich Erdbeeren oder Blaubeeren aßen, berichteten von Verbesserungen des Blutdrucks, des Gesamt- und LDL-Cholesterins und sogar von Anzeichen von oxidativem Stress. Beeren haben auch krebshemmende Eigenschaften und helfen nachweislich dabei, den mit dem Alter einhergehenden kognitiven Abbau zu verhindern.
Granatapfel
Der bekannteste sekundäre Pflanzenstoff in Granatäpfeln, Punicalagin, ist für mehr als die Hälfte der antioxidativen Aktivität der Frucht verantwortlich. Die sekundären Pflanzenstoffe des Granatapfels wirken krebsbekämpfend, kardioprotektiv und fördern die Gesundheit des Gehirns. In einer Forschungsstudie schnitten ältere Erwachsene, die 28 Tage lang täglich Granatapfelsaft tranken, bei einem Gedächtnistest besser ab als diejenigen, die ein Placebo-Getränk tranken.
Bohnen
Der Verzehr von Bohnen und anderen Hülsenfrüchten kann helfen, den Blutzuckerspiegel auszugleichen, den Appetit zu reduzieren und vor Darmkrebs zu schützen. Bohnen sind ein Anti-Diabetes-Lebensmittel, das die Gewichtsabnahme fördern kann, da sie langsam verdaut werden, was den Anstieg des Blutzuckers nach einer Mahlzeit verlangsamt und durch die Förderung des Sättigungsgefühls Heißhungerattacken vorbeugt. Es wurde festgestellt, dass der Verzehr von Bohnen und anderen Hülsenfrüchten zweimal pro Woche das Risiko für Darmkrebs senkt. Der Verzehr von Bohnen und anderen Hülsenfrüchten wie roten Bohnen, schwarzen Bohnen, Kichererbsen, Linsen und Spalterbsen bietet auch einen erheblichen Schutz vor anderen Krebsarten.
Pilze
Der regelmäßige Verzehr von Pilzen ist mit einem geringeren Brustkrebsrisiko verbunden. Weiße Pilze und Portobello-Pilze sind besonders wirksam gegen Brustkrebs, da sie Aromatasehemmer oder Verbindungen enthalten, die die Östrogenproduktion hemmen. Pilze haben nachweislich eine entzündungshemmende Wirkung und sorgen für eine verbesserte Aktivität der Immunzellen, verhindern DNA-Schäden, verlangsamen das Wachstum von Krebszellen und hemmen die Angiogenese. Pilze sollten immer gekocht werden, da rohe Pilze eine potenziell krebserregende Chemikalie namens Agaritin enthalten, die durch Kochen deutlich reduziert wird.
Zwiebeln und Knoblauch
Zwiebeln und Knoblauch wirken sich positiv auf das Herz-Kreislauf-System und das Immunsystem aus und wirken antidiabetisch und krebsbekämpfend. Diese wurden auch mit einem geringeren Risiko für Magen- und Prostatakrebs in Verbindung gebracht. Zwiebeln und Knoblauch sind für ihre Organoschwefelverbindungen bekannt, die dazu beitragen, die Entstehung von Krebs zu verhindern, indem sie Karzinogene entgiften, das Wachstum von Krebszellen verringern und die Angiogenese blockieren. Zwiebeln und Knoblauch enthalten außerdem hohe Konzentrationen an gesundheitsfördernden Flavonoid-Antioxidantien, die entzündungshemmend wirken und zur Krebsprävention beitragen können.
Tomaten
Tomaten sind reich an einer Vielzahl von Nährstoffen wie Lycopin, Vitamin C und E, Beta-Carotin und Flavonol-Antioxidantien. Lycopin kann zum Schutz vor Prostatakrebs, UV-Hautschäden usw. beitragen. Herzkreislauferkrankung. Lycopin wird beim Kochen von Tomaten besser aufgenommen. Eine Tasse Tomatensauce enthält etwa zehnmal so viel Lycopin wie eine Tasse rohe, gehackte Tomaten. Denken Sie auch daran, dass Carotinoide wie Lycopin am besten absorbiert werden, wenn sie von gesunden Fetten begleitet werden. Genießen Sie Ihre Tomaten also in einem Salat mit Nüssen oder einem Dressing auf Nussbasis für zusätzliche Nährwerte.
Die Lebensmittel, die wir essen, können unserer Gesundheit zuträglich oder schädlich sein. Eine schlechte Ernährung kann eine Vielzahl von Gesundheitsproblemen verursachen, darunter Fettleibigkeit, Herz-Kreislauf-Erkrankungen und Typ-2-Diabetes. Unterdessen kann die richtige Ernährung dazu führen, dass Sie sich energiegeladen fühlen, das Risiko von Gesundheitsproblemen verringern und dabei helfen, ein gesundes Gewicht zu halten und zu regulieren. Wenn Sie ein langes Leben fördern möchten, müssen Sie Ihren Körper mit guten Lebensmitteln versorgen. Gute Lebensmittel können auch dazu beitragen, Entzündungen zu reduzieren, die mit einer Vielzahl von Gesundheitsproblemen einhergehen, darunter Gelenkschmerzen und Arthritis. Medizinische Fachkräfte wie Chiropraktiker können Ratschläge zu Ernährung und Lebensstil geben, um Gesundheit und Wohlbefinden zu fördern. Im folgenden Artikel werden wir einige gute Lebensmittel auflisten, die letztendlich dazu beitragen können, die Langlebigkeit zu fördern. – Dr. Alex Jimenez DC, CCST Insight
Saftiger Rüben-Saft
Portionen: 1 Kochzeit: 5-10 Minuten
1 Grapefruit, geschält und in Scheiben geschnitten
1 Apfel, gewaschen und in Scheiben geschnitten
1 ganze Rübe und Blätter, wenn Sie sie haben, gewaschen und in Scheiben geschnitten
1-Zoll-Knopf Ingwer, gespült, geschält und gehackt
Alle Zutaten in einer hochwertigen Saftpresse entsaften. Am besten sofort servieren.
Nur eine Karotte gibt Ihnen die gesamte tägliche Vitamin-A-Aufnahme
Ja, wenn Sie nur eine gekochte Karotte von 80 g essen, erhalten Sie genug Beta-Carotin, damit Ihr Körper 2 Mikrogramm (mcg) Vitamin A (notwendig für die Erneuerung der Hautzellen) produziert. Das ist mehr als die empfohlene tägliche Aufnahme von Vitamin A in den USA, die etwa 1,480 mcg beträgt. Es ist am besten, gekochte Karotten zu essen, da dies die Zellwände weicher macht und mehr Beta-Carotin absorbiert werden kann. Das Hinzufügen gesünderer Lebensmittel zu Ihrer Ernährung ist eine großartige Möglichkeit, Ihre allgemeine Gesundheit zu verbessern.
Der Umfang unserer Informationen beschränkt sich auf Artikel, Themen und Diskussionen in den Bereichen Chiropraktik, Bewegungsapparat, physikalische Medizin, Wellness und sensible Gesundheit sowie auf funktionelle Medizin. Wir verwenden funktionelle Gesundheits- und Wellnessprotokolle zur Behandlung und Unterstützung der Behandlung von Verletzungen oder Störungen des Bewegungsapparates. Unsere Beiträge, Themen, Themen und Erkenntnisse decken klinische Fragen, Probleme und Themen ab, die sich direkt oder indirekt auf unseren klinischen Tätigkeitsbereich beziehen und ihn unterstützen. * Unser Büro hat einen vernünftigen Versuch unternommen, unterstützende Zitate bereitzustellen, und die relevante Forschungsstudie identifiziert oder Studien zur Unterstützung unserer Beiträge. Auf Anfrage stellen wir dem Vorstand und / oder der Öffentlichkeit auch Kopien unterstützender Forschungsstudien zur Verfügung. Wir verstehen, dass wir Angelegenheiten abdecken, die eine zusätzliche Erklärung erfordern, wie sie in einem bestimmten Pflegeplan oder Behandlungsprotokoll hilfreich sein können. Um das oben genannte Thema weiter zu diskutieren, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900. Die in Texas * & New Mexico * lizenzierten Anbieter
Kuratiert von Dr. Alex Jimenez DC, CCST
References:
Joel Fuhrman, MD. „Die 10 besten Lebensmittel, die Sie essen können, um länger zu leben und gesund zu bleiben.“ Sehr gute Gesundheit, 6. Juni 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. "Kaffee ist eine Frucht und andere unglaublich wahre Lebensmittel Fakten." MSN Lebensstil, 4. Juni 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Wenn Sie eine dieser Situationen erleben, sind Ihre Kollagenpeptide möglicherweise niedrig.
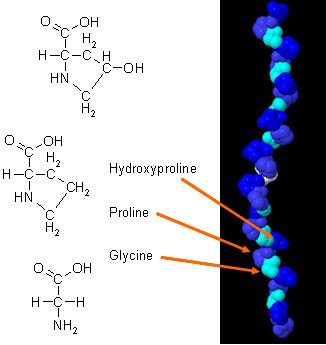
Es gibt waren neue Studien darüber, wie Kollagen die Körperzusammensetzung verbessern kann, wenn es mit täglichen Übungen kombiniert wird. Kollagen im Körper hat eine einzigartige Aminosäurezusammensetzung, die eine wesentliche Rolle in der Anatomie des Körpers spielt. Kollagenprotein ist eine konzentrierte Quelle von Glycin, Prolin und Hydroxyprolin, und wenn es mit allen anderen Nahrungsproteinen verglichen wird, macht es Kollagen zu einer möglichen praktischen Wahl als Strukturprotein.
In a 2015 Studie, haben Forscher gezeigt, wie effiziente Kollagenpräparate die Körperzusammensetzung bei aktiven Männern verbessern können. Die Ergebnisse zeigen, dass jeder Mann mindestens dreimal pro Woche am Krafttraining teilnimmt und mindestens 15 Gramm Kollagenpeptide ergänzen muss, um maximale Gesundheit zu erreichen. Die Bewertungen, die der Test bietet, sind Krafttests, Bioimpedanzanalyse (BIA) und Muskelbiopsien. Diese Tests stellen sicher, dass die männlichen Personen nach der Einnahme der Kollagenpräparate gut abschneiden, und die Ergebnisse zeigen, wie ihre Körpermasse eine Zunahme der fettfreien Körpermasse hatte. Eine andere Studie zeigte, wie eine Kollagenprotein-Supplementierung in Kombination mit einem Krafttraining die Muskelmasse und Muskelkraft bei älteren Menschen sowie bei Menschen mit Sarkopenie erhöhen kann.
Wohltuende Eigenschaften mit Kollagen
Es gibt viele vorteilhafte Eigenschaften die Kollagenergänzungen dem Körper bei der Einnahme zur Verfügung stellen können. Es gibt hydrolysiertes Kollagen und Gelatine und können helfen, die Hautstruktur einer Person zu verbessern. Auch wenn es nicht viele Studien zu Kollagenpräparaten gibt, gibt es hervorragende Versprechen für die Bereiche am Körper. Sie sind:
Muskelmasse: Kollagenpräparate können in Kombination mit Krafttraining die Muskelmasse und Kraft im Körper erhöhen.
Arthritis: Kollagenpräparate können Menschen mit Arthrose helfen. Studien zeigen dass Menschen mit Osteoarthritis, die Kollagenpräparate einnehmen, einen massiven Rückgang der Schmerzen festgestellt haben.
Hautelastizität: Im a 2014 Studie, heißt es, dass Frauen, die Kollagenpräparate einnahmen, eine Verbesserung der Hautelastizität gezeigt haben. Kollagen kann auch in topischen Behandlungen verwendet werden, um das Erscheinungsbild der Haut einer Person zu verbessern, indem feine Linien und Falten minimiert werden.
Nicht nur Kollagenpräparate bieten den spezifischen Bereichen des Körpers positive Eigenschaften, sondern es gibt die vier Haupttypen von Kollagen und ihre Rollen im menschlichen Körper sowie ihre Funktionen:
Typ 1: Kollagen vom Typ 1 macht 90% des Kollagens des Körpers aus und besteht aus dicht gepackten Fasern, die der Haut, den Knochen, dem Bindegewebe und den Zähnen des Körpers Strukturen verleihen.
Typ 2: Kollagen Typ 2 besteht aus locker gepackten Fasern, die sich im elastischen Knorpel befinden und die Gelenke im Körper polstern.
Typ 3: Kollagen Typ 3 unterstützt die Struktur der Muskeln, Organe und Arterien, die dafür sorgen, dass der Körper richtig funktioniert.
Typ 4: Kollagen Typ 4 befindet sich in den Schichten jeder Haut und hilft bei der Filtration im Körper.
Da diese vier Arten von Kollagen im Körper vorhanden sind, ist es wichtig zu wissen, dass Kollagen im Laufe der Zeit auf natürliche Weise mit dem Alter abnehmen kann, da der Körper eine geringere Kollagenqualität produziert. Eines der sichtbaren Anzeichen für eine Abnahme des Kollagens ist, wenn die Haut des menschlichen Körpers aufgrund des Alterns weniger fest und geschmeidig wird und der Knorpel schwächt.
Faktoren, die Kollagen schädigen können
Obwohl Kollagen mit zunehmendem Alter auf natürliche Weise abnehmen kann, können viele Faktoren Kollagene zerstören, die für die Haut schädlich sind. Zu den schädlichen Faktoren können gehören:
Zucker und Kohlenhydrate: Raffinierter Zucker und Kohlenhydrate stören kann mit der Fähigkeit von Kollagen, sich auf der Haut selbst zu reparieren. Durch die Minimierung des Zucker- und Kohlenhydratverbrauchs im Körper kann es die Auswirkungen von Funktionsstörungen des Gefäß-, Nieren- und Hautgewebes reduzieren.
Sonneneinstrahlung: Obwohl genug Sonne einer Person helfen kann, den Tag zu genießen, ist es jedoch für einen längeren Zeitraum der Sonne ausgesetzt kann beschädigt werden auf die Haut und zerstören Kollagenpeptide. Die Auswirkungen einer übermäßigen Sonnenbestrahlung können zu einer Lichtalterung der Haut und zu oxidativem Stress im Körper führen.
Rauchen: Wenn jemand raucht, kann das passieren die Kollagenproduktion reduzieren im Körper, was zu vorzeitigen Falten im Körper führt, und wenn der Körper verwundet ist, verlangsamt sich der Heilungsprozess und kann zu Beschwerden im Körper führen.
Autoimmunerkrankungen: Einige Autoimmunkrankheiten können auch die Kollagenproduktion wie Lupus schädigen.
Zusammenfassung
Kollagen ist für den Körper lebenswichtig, da es der Haut hilft, zart und straff zu sein. Natürlich nimmt es mit zunehmendem Alter ab, so dass die Einnahme von Kollagenpräparaten sicherstellen kann, dass der Körper richtig funktionieren kann. Wenn schädliche Faktoren auf den Körper einwirken, können sie die Kollagenproduktion stoppen oder sogar schädigen und die Bildung vorzeitiger Falten beschleunigen, wodurch eine Person älter aussieht als sie ist. Etwas Produkte kann die Zellaktivität des Körpers unterstützen, indem es eine bessere Stabilität, Bioverfügbarkeit und Verdauungskomfort bietet.
Der Umfang unserer Informationen beschränkt sich auf chiropraktische, muskuloskelettale und nervöse Gesundheitsprobleme oder Artikel, Themen und Diskussionen zur Funktionsmedizin. Wir verwenden funktionelle Gesundheitsprotokolle zur Behandlung von Verletzungen oder Erkrankungen des Bewegungsapparates. Unser Büro hat einen vernünftigen Versuch unternommen, unterstützende Zitate zur Verfügung zu stellen, und hat die relevanten Forschungsstudien oder Studien identifiziert, die unsere Stellen unterstützen. Auf Anfrage stellen wir dem Vorstand und der Öffentlichkeit auch Kopien von unterstützenden Forschungsstudien zur Verfügung. Um das Thema weiter oben zu diskutieren, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900.
References:
Bosch, Ricardo et al. Mechanismen der Photoalterung und der kutanen Photokarzinogenese und photoprotektive Strategien mit Phytochemikalien. Antioxidantien (Basel, Schweiz), MDPI, 26. März 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, F. William. Ernährung und alternde Haut: Zucker und Glykation Kliniken für Dermatologie, US National Library of Medicine, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. Kollagen – was ist es und wofür ist es gut? Healthline, 9. September 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. Neue Studie zeigt die Vorteile von Kollagenpeptiden zur Verbesserung der Körperzusammensetzung in Kombination mit Bewegung. Entwürfe für Gesundheit, 31. Mai 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. Rauchen beeinflusst die Kollagensynthese und den Umsatz der extrazellulären Matrix in der menschlichen Haut. Das British Journal of Dermatology, US National Library of Medicine, April 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E. et al. Die orale Supplementierung spezifischer Kollagenpeptide hat positive Auswirkungen auf die Physiologie der menschlichen Haut: eine doppelblinde, placebokontrollierte Studie. Hautpharmakologie und Physiologie, US National Library of Medicine, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G. et al. Wirkung des neuartigen niedermolekularen hydrolysierten Hühnersprossenknorpelextrakts, BioCell Collagen, auf die Verbesserung von Arthrose-bedingten Symptomen: eine randomisierte, doppelblinde, placebokontrollierte Studie. Zeitschrift für Agrar- und Lebensmittelchemie, US-amerikanische Nationalbibliothek für Medizin, 25 Apr. 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise et al. Kollagen-Peptid-Supplementierung in Kombination mit Krafttraining verbessert die Körperzusammensetzung und erhöht die Muskelkraft bei älteren Männern mit Sarkopenie: eine randomisierte kontrollierte Studie. Das britische Journal der Ernährung, Cambridge University Press, 28. Oktober 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Indem sie Einzelpersonen darüber informiert, wie die National University of Health Sciences das Wissen für zukünftige Generationen bereitstellt, bietet die Universität eine Vielzahl von medizinischen Berufen für die funktionelle Medizin an.
Als ob bei Ihnen Zöliakie, Reizdarmsyndrom, Divertikulose/Divertikulitis oder Leaky-Gut-Syndrom diagnostiziert worden wäre?
Übermäßiges Aufstoßen, Aufstoßen oder Blähungen?
Abnormale Blähungen nach bestimmten Probiotika oder natürlichen Nahrungsergänzungsmitteln?
Verdacht auf ernährungsbedingte Malabsorption?
Lassen Verdauungsprobleme durch Entspannung nach?
Wenn eine dieser Situationen bei Ihnen auftritt, haben Sie möglicherweise Darmprobleme und müssen möglicherweise das 4R-Programm ausprobieren.
Nahrungsmittelunverträglichkeiten, rheumatoide Arthritis und Angstzustände wurden mit einer beeinträchtigten Magen-Darm-Permeabilität in Verbindung gebracht. Diese verschiedenen Erkrankungen können durch viele Faktoren verursacht werden, die sich auf den Verdauungstrakt auswirken können. Wenn es unbehandelt bleibt, kann es möglicherweise die Folge einer Funktionsstörung der Darmpermeabilitätsbarriere sein, die zu Entzündungen und schwerwiegenden Gesundheitszuständen im Darm führen kann. Das 4R-Programm dient der Wiederherstellung eines gesunden Darms im Körper und umfasst vier Schritte. Dies sind: Entfernen, Ersetzen, Neuimpfen und Reparieren.
Darmdurchlässigkeit
Die Darmdurchlässigkeit schützt den Körper und sorgt dafür, dass keine schädlichen Bakterien in den Darm gelangen. Es schützt den Körper vor mögliche Umweltfaktoren die schädlich sein können und über den Verdauungstrakt gelangen. Dabei kann es sich entweder um Toxine, pathogene Mikroorganismen oder andere Antigene handeln, die den Verdauungstrakt schädigen und Probleme verursachen können. Die Darmschleimhaut besteht aus einer Schicht Epithelzellen, die durch enge Verbindungen voneinander getrennt sind. In einem gesunden DarmDie Tight Junction reguliert die Darmpermeabilität, indem sie Substanzen selektiv in die Darmbarriere eindringen und diese passieren lässt und verhindert, dass schädliche Faktoren absorbiert werden.
Bestimmte Umweltfaktoren können die enge Verbindung schädigen und dadurch die Darmpermeabilität erhöhen, was zu einer intestinalen Hyperpermeabilität oder einem Leaky Gut im Körper führt. Beteiligte Faktoren wie eine übermäßige Menge an gesättigten Fetten und Alkohol, Nährstoffmangel, chronischer Stress und Infektionskrankheiten können die Darmpermeabilität erhöhen.
Mit einer erhöhten Darmpermeabilität Im Darm kann es Antigenen ermöglichen, die Darmschleimhaut zu passieren und in den Blutkreislauf zu gelangen, wodurch eine Immunantwort und eine Entzündung im Körper ausgelöst wird. Es gibt bestimmte Magen-Darm-Erkrankungen, die mit einer Überpermeabilität des Darms einhergehen und unbehandelt bestimmte Autoimmunerkrankungen auslösen können, die dem Körper schaden können.
4Rs-Programm
Bei den 4Rs handelt es sich um ein Programm, das medizinische Fachkräfte ihren Patienten empfehlen, wenn sie störende Verdauungsprobleme angehen und die Darmheilung unterstützen.
Beheben des Problems
Der erste Schritt im 4R-Programm besteht darin, schädliche Krankheitserreger und Entzündungsauslöser zu entfernen, die mit einer erhöhten Darmpermeabilität verbunden sind. Auslöser wie Stress und chronischer Alkoholkonsum können dem Körper eines Menschen großen Schaden zufügen. Um diese schädlichen Faktoren aus dem Körper zu entfernen, müssen Sie ihn mit Medikamenten, Antibiotika und Nahrungsergänzungsmitteln behandeln. Außerdem wird empfohlen, entzündungsfördernde Lebensmittel aus der Ernährung zu streichen, darunter:
- Alkohol
- Gluten
- Lebensmittelzusatzstoffe
– Stärken
– Bestimmte Fettsäuren
– Bestimmte Lebensmittel, auf die eine Person empfindlich reagiert
Ersetzen der Nährstoffe
Der zweite Schritt des 4R-Programms besteht darin, die Nährstoffe zu ersetzen, die durch Entzündungen die Darmprobleme verursachen. Bestimmte Nährstoffe können dazu beitragen, Entzündungen im Darm zu reduzieren und gleichzeitig dafür zu sorgen, dass der Verdauungstrakt unterstützt wird. Es gibt einige entzündungshemmende Lebensmittel, die nahrhaft sind. Diese beinhalten:
– Ballaststoffreiche Lebensmittel
– Omega-3-Fettsäuren
- Olivenöl
- Pilze
– Entzündungshemmende Kräuter
Es gibt bestimmte Nahrungsergänzungsmittel, die zur Unterstützung der Verdauungsfunktion verwendet werden können, indem sie die Nährstoffe unterstützen und absorbieren, um einen gesunden Darm zu fördern. Die Aufgabe der Verdauungsenzyme besteht darin, dass sie beim Abbau von Fetten, Proteinen und Kohlenhydraten im Darm helfen. Dies kommt Personen zugute, die unter einer Beeinträchtigung des Verdauungstrakts, Nahrungsmittelunverträglichkeiten oder Zöliakie leiden. Nahrungsergänzungsmittel wie Gallensäurepräparate können die Nährstoffaufnahme unterstützen, indem sie Lipide miteinander verbinden. Studien haben ergeben dass Gallensäuren zur Behandlung von Leber, Gallenblase und Gallengang eingesetzt wurden und gleichzeitig die Bildung von Gallensteinen nach einer bariatrischen Operation verhindert wurden.
Den Darm neu geimpft
Der dritte Schritt besteht im Rahmen des 4rs-Programms darin, die Darmmikroben mit nützlichen Bakterien neu zu inokulieren, um eine gesunde Darmfunktion zu fördern. Studien wurden gezeigt dass probiotische Nahrungsergänzungsmittel verwendet wurden, um den Darm durch die Wiederherstellung nützlicher Bakterien zu verbessern. Mit diesen Nahrungsergänzungsmitteln verbessern sie den Darm, indem sie entzündungshemmende Substanzen in den Körper abgeben, das Immunsystem unterstützen, die mikrobielle Zusammensetzung des Körpers verändern und die Darmpermeabilität im Darmsystem verringern.
Da Probiotika werden gefunden in fermentierten Lebensmitteln und gelten als vorübergehend, da sie im Magen-Darm-Trakt nicht persistieren und von Nutzen sind. Überraschenderweise haben sie immer noch einen Einfluss auf die menschliche Gesundheit, da sie den Darm durch die Produktion von Vitaminen und antimikrobiellen Verbindungen beeinflussen und so für Vielfalt und Darmfunktion sorgen.
Den Darm reparieren
Der letzte Schritt des 4R-Programms besteht darin, den Darm zu reparieren. Bei diesem Schritt wird die Darmschleimhaut mit spezifischen Nährstoffen und Kräutern repariert. Diese Kräuter und Nahrungsergänzungsmittel können dazu beitragen, die Darmpermeabilität und Entzündungen im Körper zu verringern. Zu diesen Kräutern und Nahrungsergänzungsmitteln gehören:
- Aloe Vera
– Chios-Mastixgummi
– DGL (deglycyrrhiziniertes Lakritz)
- Marshmallowwurzel
– L-Glutamin
– Omega-3-Fettsäuren
� Polyphenole
- Vitamin-D
– Zink
Zusammenfassung
Denn viele Faktoren können das Verdauungssystem auf schädliche Weise beeinträchtigen und zu verschiedenen Gesundheitszuständen beitragen. Das Hauptziel des 4R-Programms besteht darin, diese Faktoren, die den Darm schädigen, zu minimieren, Entzündungen zu reduzieren und die Darmpermeabilität zu erhöhen. Wenn der Patient mit den positiven Faktoren vertraut gemacht wird, die die 4R bieten, kann dies zu einem gesunden, geheilten Darm führen. Manche Produkte sind dazu da, das Magen-Darm-System zu unterstützen, indem sie den Darm unterstützen, den Zuckerstoffwechsel verbessern und auf die Aminosäuren abzielen, die den Darm unterstützen sollen.
Der Umfang unserer Informationen beschränkt sich auf chiropraktische, muskuloskelettale und nervöse Gesundheitsprobleme oder Artikel, Themen und Diskussionen zur Funktionsmedizin. Wir verwenden funktionelle Gesundheitsprotokolle zur Behandlung von Verletzungen oder Erkrankungen des Bewegungsapparates. Unser Büro hat einen vernünftigen Versuch unternommen, unterstützende Zitate zur Verfügung zu stellen, und hat die relevanten Forschungsstudien oder Studien identifiziert, die unsere Stellen unterstützen. Auf Anfrage stellen wir dem Vorstand und der Öffentlichkeit auch Kopien von unterstützenden Forschungsstudien zur Verfügung. Um das Thema weiter oben zu diskutieren, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900.
References:
De Santis, Stefania et al. „Ernährungsschlüssel für die Modulation der Darmbarriere.“ Grenzen in der Immunologie, Frontiers Media SA, 7 Dez. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca et al. „Verdauungsenzym-Supplementierung bei Magen-Darm-Erkrankungen.“ Aktueller Drogenstoffwechsel, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui et al. „Leaky Gut als Gefahrensignal für Autoimmunerkrankungen.“ Frontiers, Frontiers, 5. Mai 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon et al. „Fermentierte Lebensmittel als Nahrungsquelle für lebende Organismen.“ Grenzen in der Mikrobiologie, Frontiers Media SA, 24. August 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., et al. „Eine schnelle Störung der Darmbarrierefunktion durch Gliadin führt zu einer veränderten Expression apikaler Verbindungsproteine.“ FEBS-Presse, John Wiley & Sons, Ltd, 8. August 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R. Balfour. „Therapeutische Manipulation der enterischen Mikroflora bei entzündlichen Darmerkrankungen: Antibiotika, Probiotika und Präbiotika.“ Gastroenterology, US National Library of Medicine, Mai 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Chronische Schmerzen sind ein weit verbreitetes Gesundheitsproblem, von dem viele Menschen in den Vereinigten Staaten betroffen sind. Während verschiedene Erkrankungen, wie Fibromyalgie und myofasziale Schmerzsyndrom, chronische Schmerzen verursachen können, können sie sich auch aufgrund einer Vielzahl anderer gesundheitlicher Probleme entwickeln. Forschungsstudien haben ergeben, dass weit verbreitete Entzündungen die Hauptursache für chronische Schmerzen sind. Entzündungen sind ein natürlicher Abwehrmechanismus gegen Verletzungen, Krankheiten oder Infektionen. Wenn der Entzündungsprozess jedoch zu lange andauert, kann er problematisch werden.
Entzündungen signalisieren dem Immunsystem, geschädigtes Gewebe zu heilen und zu reparieren sowie sich vor Bakterien und Viren zu schützen. Wie bereits erwähnt, kann eine chronische Entzündung jedoch eine Reihe von gesundheitlichen Problemen verursachen, einschließlich chronischer Schmerzsymptome. Ein gesunder Lebensstil kann dazu beitragen, chronische Schmerzen zu behandeln, aber zunächst sollten wir die häufigsten Ursachen für chronische Schmerzen verstehen.
Was ist eine akute Entzündung?
Eine akute Entzündung tritt zum Beispiel nach einer Verletzung oder so etwas wie Halsschmerzen auf. Es ist eine natürliche Reaktion mit negativen Auswirkungen, was bedeutet, dass es lokal in der Region wirkt, in der das Gesundheitsproblem gefunden wird. Die häufigsten Anzeichen für eine akute Entzündung sind Schwellungen, Rötungen, Wärme, Schmerzen und Funktionsverlust, wie von der National Library of Medicine festgestellt. Wenn sich eine akute Entzündung entwickelt, dehnen sich die Blutgefäße aus, was zu einer Erhöhung des Blutflusses führt, und weiße Blutkörperchen in der verletzten Region fördern die Erholung.
Bei einer schweren Entzündung werden Verbindungen, die als Zytokine bezeichnet werden, durch das beschädigte Gewebe freigesetzt. Die Zytokine fungieren als „Notfallsignale“, die die körpereigenen Immunzellen sowie Hormone und zahlreiche Nährstoffe zur Behebung des Gesundheitsproblems anregen. Außerdem verursachen hormonähnliche Substanzen, so genannte Prostaglandine, Blutgerinnsel, die das geschädigte Gewebe heilen, und diese können im Rahmen der Entzündungsprozedur auch Fieber und Schmerzen auslösen. Sobald sich der Schaden oder die Verletzung erholt, lässt die Entzündung nach.
Was ist eine chronische Entzündung?
Im Gegensatz zu akuten Entzündungen hat chronische Entzündung Langzeiteffekte. Chronische Entzündungen, auch als persistierende Entzündungen bezeichnet, führen im gesamten menschlichen Körper zu einer niedrigen Entzündungsrate, was durch eine Zunahme der Marker des Immunsystems im Blut und im Zellgewebe gezeigt wird. Chronische Entzündungen können auch das Fortschreiten verschiedener Krankheiten und Zustände verursachen. In manchen Fällen kann es zu einer erhöhten Entzündung kommen, auch wenn keine Verletzungen, Krankheiten oder Infektionen vorliegen, die auch das Immunsystem ansprechen können.
Infolgedessen könnte das Immunsystem des menschlichen Körpers beginnen, gesunde Zellen, Gewebe oder Organe anzugreifen. Forscher versuchen immer noch, die Folgen chronischer Entzündungen im menschlichen Körper und die Mechanismen dieses natürlichen Abwehrprozesses zu verstehen. Chronische Entzündungen wurden beispielsweise mit einer Vielzahl von Gesundheitsproblemen wie Herzerkrankungen und Schlaganfällen in Verbindung gebracht.
Eine Theorie besagt, dass, wenn eine Entzündung in den Blutgefäßen verbleibt, dies die Ansammlung von Plaque fördern kann. Laut der American Heart Association oder der AHA können die weißen Blutkörperchen, wenn das Immunsystem Plaque als einen fremden Eindringling identifiziert, versuchen, sich von der Plaque abzuwenden, die im Blut gefunden wird, das durch die Arterien fließt. Dadurch kann ein Blutgerinnsel entstehen, das den Blutfluss zum Herzen oder zum Gehirn blockieren kann, wodurch es instabil wird und reißt. Krebs ist ein weiteres Gesundheitsproblem, das mit chronischen Entzündungen einhergeht. Laut dem National Cancer Institute können DNA-Schäden auch durch chronische Entzündungen verursacht werden.
Bei anhaltenden, minderwertigen Entzündungen treten häufig keine Symptome auf. Die Angehörigen der Gesundheitsberufe können jedoch nach einem C-reaktiven Protein oder CRP (Liponsäure) suchen, einem Marker für Entzündungen im Blut. Erhöhte CRP-Spiegel gehen mit einem erhöhten Risiko für Herz-Kreislauf-Erkrankungen einher. Erhöhte CRP-Spiegel können bei chronischen Erkrankungen wie Lupus oder rheumatoider Arthritis auftreten.
Bei anderen chronischen Erkrankungen, wie z. B. Fibromyalgie, reagiert das Nervensystem auf eine bestimmte Stimulation, die Entzündung führt jedoch zu chronischen Schmerzsymptomen. Subjektiv ist es fast unmöglich, den Unterschied zwischen chronischen Schmerzen, die durch ein überempfindliches Nervensystem verursacht werden, und chronischen Schmerzen, die durch weit verbreitete Entzündungen verursacht werden, zu erkennen. Neben der Suche nach Hinweisen im Blutkreislauf können Ernährung, Lebensgewohnheiten und Umwelteinflüsse einer Person auch chronische Entzündungen fördern.
Eine Entzündung ist der natürliche Abwehrmechanismus des Immunsystems gegen Verletzungen, Krankheiten oder Infektionen. Während diese Entzündungsreaktion zur Heilung und Reparatur von Geweben beitragen kann, kann eine chronische, weit verbreitete Entzündung eine Vielzahl von Gesundheitsproblemen verursachen, einschließlich chronischer Schmerzsymptome. Ein ausgeglichenes Ernährung, einschließlich einer Vielzahl von Diäten und Fasten, kann helfen, Entzündungen zu reduzieren. Das Fasten, auch als Kalorienrestriktion bekannt, fördert die Zellapoptose und die mitochondriale Erholung. Die Fastennachahmungsdiät, die Teil des Langlebigkeitsdiätplans ist, ist ein Diätprogramm, das den menschlichen Körper in einen Fastenzustand versetzt, um die Vorteile des traditionellen Fastens zu erfahren. Konsultieren Sie unbedingt einen Arzt, bevor Sie eine der in diesem Artikel beschriebenen Diäten befolgen.
Dr. Alex Jiménez DC, CCST Insight
Ernährung, Diäten, Fasten und chronische Schmerzen
Entzündungshemmende Diäten bestehen hauptsächlich aus frischem Obst und Gemüse, Fisch und Fetten. Bei der Mittelmeerdiät handelt es sich beispielsweise um eine entzündungshemmende Diät, die den Verzehr mäßiger Mengen an Nüssen, das Einnehmen von sehr wenig Fleisch und das Trinken von Wein fördert. Entzündungshemmende Nahrungsbestandteile, wie Omega-3-Fettsäuren, schützen den menschlichen Körper vor der Haut daMagier durch Entzündung hervorgerufen.
Eine entzündungshemmende Diät beinhaltet auch, sich von Nahrungsmitteln fernzuhalten, die Entzündungen fördern können. Es ist ideal, um die Menge an Lebensmitteln, die Sie essen, zu reduzieren, die reich an trans- und gesättigten Fetten sind, wie z. B. Fleisch. Darüber hinaus begrenzt eine entzündungshemmende Diät den Verbrauch von raffinierten Kohlenhydraten und Lebensmitteln wie Brot und Reis. Dies trägt auch dazu bei, die Verwendung von Margarine und Ölen, die mit Omega-6-Fettsäuren gefüllt sind, wie Sonnenblumen, Saflor, zu reduzieren und Maisöle.
Es ist seit langem bekannt, dass Fasten oder kalorische Einschränkung oxidativen Stress verringert und die Alterungsmechanismen in verschiedenen Organismen verlangsamt. Die Auswirkungen des Fastens umfassen den programmierten Zelltod oder Apoptose, Transkription, mobile Energieeffizienz, mitochondriale Biogenese, antioxidative Mechanismen und den zirkadianen Rhythmus. Das Fasten trägt auch zur mitochondrialen Autophagie bei, der sogenannten Mitophagie, bei der Gene in den Mitochondrien zur Apoptose stimuliert werden, was die Mitochondrien-Genesung fördert.
Intermittierendes Fasten kann Ihnen helfen, Entzündungen zu bekämpfen, die Verdauung zu verbessern und Ihre Lebenserwartung zu steigern. Der menschliche Körper ist so konzipiert, dass er längere Zeit ohne Nahrung überleben kann. Forschungsstudien haben gezeigt, dass intermittierendes Fasten positive Veränderungen in der Gesamtzusammensetzung Ihrer Darmmikrobiota bewirken kann. Darüber hinaus kann intermittierendes Fasten die Insulinresistenz verringern und gleichzeitig die Reaktion des Immunsystems erhöhen. Schließlich kann intermittierendes Fasten die Produktion einer Substanz fördern, die als & agr; -Hydroxybutyrat bekannt ist und einen Teil des Immunsystems blockiert, das an entzündlichen Erkrankungen beteiligt ist, sowie die Produktion von Entzündungsmarkern wie Zytokinen und dem C-reaktiven Protein erheblich verringern oder CRP, zuvor erwähnt.
Der in dem Buch von Dr. Valter Longo vorgestellte Langlebigkeitsdiätplan beseitigt den Verzehr von verarbeiteten Lebensmitteln, die Entzündungen verursachen können und das Wohlbefinden und die Langlebigkeit fördern. Dieses einzigartige Ernährungsprogramm fördert im Gegensatz zu den meisten traditionellen Diäten den Gewichtsverlust nicht. Obwohl Sie möglicherweise an Gewicht verlieren, liegt der Schwerpunkt dieses einzigartigen Ernährungsprogramms auf dem gesünderen Essen. Es wurde gezeigt, dass der Longevity-Diätplan dazu beiträgt, eine Stammzellen-basierte Erneuerung zu aktivieren, das Bauchfett zu reduzieren und altersbedingten Knochen- und Muskelverlust zu verhindern sowie eine Resistenz gegen Herz-Kreislauf-Erkrankungen, Alzheimer, Diabetes und Krebs aufzubauen.
Die Fasten-Nachahmungsdiät (FMD) ermöglicht Ihnen, die Vorteile des traditionellen Fastens zu erleben, ohne Ihrem Körper die Nahrung zu nehmen. Der Hauptunterschied der MKS besteht darin, dass Sie die gesamte Kalorienzufuhr für mehrere Tage oder Wochen nicht vollständig ausschließen, sondern nur fünf Tage im Monat die Kalorienzufuhr einschränken. Die MKS kann einmal im Monat geübt werden, um die allgemeine Gesundheit und das Wohlbefinden zu fördern.
Während jedermann der FMD alleine folgen kann, kann die ProLon Die Diät für das Fastennachahmungsprogramm bietet ein 5-Tagesmahlzeitprogramm, das für jeden Tag einzeln verpackt und beschriftet wurde. Es serviert die Nahrungsmittel, die Sie für die MKS benötigen, in genauen Mengen und Kombinationen. Das Mahlzeitprogramm besteht aus verzehrfertigen oder einfach zuzubereitenden, pflanzlichen Lebensmitteln, darunter Riegeln, Suppen, Snacks, Ergänzungsmitteln, einem Getränkekonzentrat und Tees. Vor dem Start des ProLon® Fasten imitiert Diät, 5-tägiges EssensprogrammStellen Sie sicher, dass Sie mit einem Arzt sprechen, um herauszufinden, welche Behandlung für chronische Schmerzen die richtige für Sie ist.
Der Umfang unserer Informationen beschränkt sich auf Artikel, Themen und Diskussionen zu Chiropraktik, Fragen der Wirbelsäulengesundheit und funktioneller Medizin. Um das oben genannte Thema weiter zu besprechen, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900 .
Rückenschmerzen ist eine der häufigsten Ursachen für Behinderungen und versäumte Arbeitstage weltweit. Rückenschmerzen sind der zweithäufigste Grund für Arztbesuche, die nur bei Infektionen der oberen Atemwege unterlegen sind. Ungefähr 80 Prozent der Bevölkerung wird mindestens einmal im Leben Rückenschmerzen haben. Ihre Wirbelsäule ist eine komplexe Struktur, bestehend aus Knochen, Gelenken, Bändern und Muskeln sowie anderen Weichteilen. Verletzungen und / oder erschwerte Bedingungen, wie z Bandscheibenvorfall, kann schließlich zu Rückenschmerzen führen. Sportverletzungen oder Autounfallverletzungen sind oft die häufigste Ursache von Rückenschmerzen, jedoch können manchmal die einfachsten Bewegungen schmerzhafte Folgen haben. Glücklicherweise können alternative Behandlungsmöglichkeiten wie Chiropraktik helfen, Rückenschmerzen durch die Verwendung von Wirbelsäuleneinstellungen und manuellen Manipulationen zu lindern, was letztlich die Schmerzlinderung verbessert.
XYMOGEN s Exklusive professionelle Formeln sind über ausgewählte, lizenzierte Fachkräfte im Gesundheitswesen erhältlich. Der Verkauf und die Diskontierung von XYMOGEN-Formeln im Internet sind strengstens untersagt.
Stolz, Dr. Alexander Jimenez macht XYMOGEN-Formeln nur für Patienten verfügbar, die unter unserer Obhut stehen.
Bitte rufen Sie unser Büro an, um eine ärztliche Konsultation für den sofortigen Zugang zu erhalten.
Wenn Sie ein Patient von sind Injury Medical & Chiropractic Clinickönnen Sie sich über XYMOGEN anrufen 915-850-0900.
Für Ihre Bequemlichkeit und Überprüfung der XYMOGEN Produkte bitte den folgenden Link überprüfen. *XYMOGEN-Katalog-Herunterladen
* Alle oben genannten XYMOGEN-Richtlinien bleiben strikt in Kraft.
Das Festhalten an einer bestimmten Diät zur Aufrechterhaltung der richtigen Ernährung kann das Essen manchmal stressig machen. Natürliche Veränderungen des Lebensstils sind der Schlüssel zur Veränderung Ihrer Essgewohnheiten. Dies kann Ihnen helfen, ein längeres und gesünderes Leben zu führen. Der von Dr. Valter Longo entwickelte Langlebigkeits-Diätplan ist eine Auswahl praktischer Essensrichtlinien, die sich darauf konzentrieren, Ihre Ernährungsmuster zu ändern, um Gesundheit und Wohlbefinden zu erreichen.
Die Regeln des Langlebigkeits-Diätplans
Indem Sie lediglich die folgenden Ernährungstipps befolgen, können Sie Ihre derzeitige Diät überholen und gesünder essen, ohne den Stress einer traditionellen Diät. Der Langlebigkeits-Diätplan beseitigt den Verzehr von verarbeiteten Lebensmitteln, die eine Vielzahl gesundheitlicher Probleme verursachen können, und erhöht den Verbrauch von Nährstoffen, die die Langlebigkeit fördern. Dieses einzigartige Ernährungsprogramm teilt die Ergebnisse von etwa 25-Jahren von Forschungsstudien mit einer einfachen Lösung, die den Menschen dabei helfen kann, durch richtige Ernährung ein allgemeines Wohlbefinden zu erzielen.
Im Gegensatz zu den meisten herkömmlichen Diäten fördert der Langlebigkeits-Diätplan jedoch nicht den Gewichtsverlust. Obwohl Sie möglicherweise eine Gewichtsreduktion erleben, liegt der Schwerpunkt dieses einzigartigen Diätprogramms auf einer gesünderen Ernährung. Es wurde gezeigt, dass der Langlebigkeits-Diätplan Ihnen dabei hilft, die Stammzellen-basierte Erneuerung zu aktivieren, Gewicht zu verlieren und Bauchfett zu reduzieren, altersbedingten Knochen- und Muskelverlust zu verhindern, Resistenzen gegen Herz-Kreislauf-Erkrankungen, Alzheimer-Krankheit, Diabetes und Krebs aufzubauen als verlängern Langlebigkeit. Im Folgenden werden die 8 häufigsten Ernährungstipps des Langlebigkeits-Diätplans zusammengefasst, die letztendlich dazu beitragen können, Ihr Leben länger und gesünder zu gestalten.
Der Langlebigkeits-Diätplan ist ein einzigartiges Diätprogramm, das von Dr. Valter Longo zur Förderung der allgemeinen Gesundheit, des Wohlbefindens und der Langlebigkeit entwickelt wurde. Durch einfache Änderungen am Lebensstil können Menschen ihre Essgewohnheiten ändern und die vielen gesundheitlichen Vorteile dieses Ernährungsprogramms nutzen. Durch eine pescatarische Diät und die Befolgung der ProLon Fasten imitiert DiätUnter den anderen unten beschriebenen Ernährungstipps können die Menschen länger und gesünder leben. Traditionelle Diäten können oft schwierig und anstrengend sein, jedoch ist der Longevity Diet Plan ein praktisches und einzigartiges Diätprogramm, das für viele Menschen geeignet sein kann.
Dr. Alex Jiménez DC, CCST Insight
8-Ernährungstipps für den Langlebigkeits-Diätplan
Befolgen Sie eine pescatarische Diät
Als Teil des Longevity-Diätplans folgen Sie einer pescatarischen Diät, die fast 100 Prozent pflanzlich und fischabhängig ist. Achten Sie außerdem darauf, den Fischkonsum auf zwei bis drei Portionen pro Woche zu beschränken, um Fische mit einem höheren Quecksilbergehalt wie Thunfisch, Schwertfisch, Makrele und Heilbutt zu vermeiden. Wenn Sie 65 überstanden haben und weniger Muskelmasse, Kraft und Fett zu spüren bekommen, fügen Sie Ihrer Ernährung neben anderen tierischen Lebensmitteln, einschließlich Eiern und bestimmten Käsesorten wie Feta oder Pecorino, und Joghurt aus Ziegenfleisch weitere Nahrung hinzu Milch.
Essen Sie nicht zu viel Protein
Gemäß dem Longevity Diet Plan sollten wir täglich 0.31 bis 0.36 Gramm Protein pro Pfund Körperfett zu uns nehmen. Wenn Sie 130 Pfund wiegen, sollten Sie etwa 40 bis 47 Gramm Protein pro . zu sich nehmen Tag oder ein Äquivalent von 1.5 Lachsfilets, 1 Tasse Kichererbsen oder 2 1/2 Tassen Linsen, von denen 30 Gramm in einer Mahlzeit verzehrt werden sollten. Wenn Sie 200 bis 220 Pfund wiegen, sollten Sie etwa 60 bis 70 Gramm Protein pro Tag essen, oder das Äquivalent von zwei Lachsfilets, 3 1/2 Tassen Linsen oder 1 1/2 Tassen Kichererbsen. Ab dem 65. Lebensjahr sollte der Proteinkonsum gesteigert werden. Für die meisten von uns reichen 10 bis 20 Prozent oder 5 bis 10 Gramm mehr pro Tag aus. Schließlich ist die Longevity Diet frei von tierischen Proteinen wie rotem Fleisch, weißem Fleisch und Geflügel, mit Ausnahme von tierischen Proteinen in Fisch. Dieses einzigartige Diätprogramm ist stattdessen vergleichsweise reich an pflanzlichen Proteinen wie Hülsenfrüchten und Nüssen, um Gesundheit und Wohlbefinden zu optimieren.
Erhöhen Sie gute Fette und komplexe Kohlenhydrate
Als Teil des Langlebigkeits-Diätplans sollten Sie höhere Mengen an mehrfach ungesättigten Fetten essen, wie sie beispielsweise in Lachs, Mandeln, Walnüssen und Olivenöl vorkommen, während Sie weniger gesättigte, hydrierte und Transfette essen sollten. Ebenso sollten Sie im Rahmen des Longevity-Diätplans auch komplexe Kohlenhydrate wie Vollkornbrot, Hülsenfrüchte und Gemüse zu sich nehmen. Vermeiden Sie es, Nudeln, Reis, Brot, Obst und Fruchtsäfte zu essen, die in Zucker umgewandelt werden können, wenn sie Ihren Darm erreichen.
Nehmen Sie Nahrungsergänzungsmittel
Der menschliche Körper benötigt Proteine, essentielle Fettsäuren wie Omega-3 und Omega-6, Vitamine, Mineralien und sogar Zucker, um richtig zu funktionieren. Wenn die Aufnahme bestimmter Nährstoffe zu niedrig ist, können sich die Reparatur-, Ersatz- und Abwehrmethoden des menschlichen Körpers verlangsamen oder aufhören, wodurch Pilze, Bakterien und Viren Schäden verursachen können, die zu einer Vielzahl von Gesundheitsproblemen führen können. Nehmen Sie Vitamin- und Mineralstoff-Nahrungsergänzungsmittel ein, insbesondere für Omega-3, wie von Ihrem Arzt empfohlen.
Essen Sie verschiedene Lebensmittel aus Ihrem AAbstammung
Um alle notwendigen Nährstoffe zu sich zu nehmen, müssen Sie eine große Auswahl an Speisen zu sich nehmen. Am besten sollten Sie jedoch Speisen wählen, die auf dem Tisch Ihrer Eltern, Großeltern und Urgroßeltern üblich waren. Beispielsweise ist in vielen nordeuropäischen Ländern, in denen Milch im Allgemeinen konsumiert wurde, die Laktoseintoleranz relativ selten, wohingegen die Laktoseintoleranz in südeuropäischen und asiatischen Ländern recht häufig ist, wo Milch traditionell nicht zur herkömmlichen Ernährung von Erwachsenen gehörte. Wenn eine Person japanischer Abstammung, die in den USA lebt, plötzlich beschließt, Milch zu trinken, die wahrscheinlich selten am Esstisch ihrer Großeltern serviert wurde, wird sie sich wahrscheinlich krank fühlen. Die häufigsten Probleme in diesen Fällen sind Unverträglichkeiten oder Autoimmunitäten, wie z. B. die Reaktion auf glutenreiche Lebensmittel wie Brot und Nudeln, die bei Zöliakie-Betroffenen auftreten. Obwohl weitere Beweise erforderlich sind, ist es möglich, dass Nahrungsmittelunverträglichkeiten mit vielen Autoimmunerkrankungen zusammenhängen, einschließlich Diabetes, Colitis und Morbus Crohn.
Essen Sie zwei Mahlzeiten pro Tag und einen Snack
Gemäß dem Langlebigkeits-Diätplan ist es ideal, jeden Tag Frühstück und eine Hauptmahlzeit sowie einen nahrhaften kalorienarmen, zuckerarmen Snack zu sich zu nehmen. Während für einige Leute empfohlen werden kann, drei Mahlzeiten und einen Snack jeden Tag zu essen. Viele Ernährungsrichtlinien empfehlen, täglich fünf bis sechs Mahlzeiten zu sich zu nehmen. Wenn Menschen geraten wird, häufig zu essen, kann es für sie oft schwierig werden, ihre Kalorienaufnahme zu regulieren. In den letzten zwanzig Jahren gelten ungefähr 70 Prozent der Bevölkerung in den Vereinigten Staaten als übergewichtig oder fettleibig. Es ist viel schwieriger, den Langlebigkeits-Diätplan zu überfressen, wenn Sie jeden Tag nur zweieinhalb Mahlzeiten zu sich nehmen. Es würde massive Portionen von Hülsenfrüchten, Gemüse und Fisch erfordern, um die Menge zu erreichen, die zu einer Gewichtszunahme führen würde. Die hohe Ernährung der Mahlzeiten sowie die Menge der Mahlzeit senden ein Signal an Ihren Magen und Ihr Gehirn, dass Sie genug zu essen hatten. Dieses eine Hauptmahlzeitsystem muss manchmal in zwei Mahlzeiten unterteilt werden, um Verdauungsprobleme zu vermeiden. Erwachsene und ältere Menschen, die zur Gewichtsreduktion neigen, sollten drei Mahlzeiten pro Tag zu sich nehmen. Für Menschen, die versuchen, Gewicht zu verlieren, sowie für Menschen, die übergewichtig oder fettleibig sind, wäre der beste Ernährungsrat, täglich zu frühstücken. essen oder zu Mittag essen, aber nicht beides, und ersetzen Sie die versäumte Mahlzeit durch einen Snack, der weniger als 100 Kalorien und nicht mehr als 3 bis 5 g Zucker enthält. Welche Mahlzeit Sie auslassen, hängt von Ihrem Lebensstil ab. Es wird jedoch aufgrund gesundheitsschädlicher Probleme nicht empfohlen, das Frühstück auszulassen. Der Vorteil des Auslassens des Mittagessens ist mehr Freizeit und Energie. Es gibt jedoch einen Nachteil beim Essen eines großen Abendessens, insbesondere für Menschen, die unter saurem Reflux oder Schlafstörungen leiden. Der Nachteil beim Auslassen des Abendessens besteht jedoch darin, dass dadurch möglicherweise die gesellschaftliche Mahlzeit ihres Tages beseitigt wird.
Essen Sie jeden Tag in einem 12-Stundenfenster
Eine andere übliche Essgewohnheit, die von vielen Hundertjährigen angenommen wird, ist das zeitabhängige Essen oder Einschränken aller Mahlzeiten und Snacks innerhalb eines 12-Stundenfensters jeden Tag. Die Wirksamkeit dieser Methode wurde sowohl in Humanstudien als auch in Tierversuchen nachgewiesen. Normalerweise frühstückte man bei 8 am und dann um 8 pm. Ein kürzeres Esszeitfenster von zehn Stunden oder weniger kann für die Gewichtsabnahme sogar noch besser sein, es ist jedoch erheblich schwieriger zu pflegen und kann das Risiko erhöhen, Nebenwirkungen wie Gallensteine zu entwickeln, und möglicherweise sogar die Wahrscheinlichkeit, an Herz-Kreislauf-Erkrankungen zu erkranken. Sie sollten nicht drei bis vier Stunden vor dem Schlafengehen essen.
Befolgen Sie die ProLon Fasten imitierende Diät
Gesunde Menschen unter 65 sollten dem folgen ProLon Fasten imitierende Diät, 5-Tage-Mahlzeitprogramm mindestens zweimal pro Jahr. Die MKS ist eines der wichtigsten Prinzipien des Longevity-Diätplans. Die Fastennachahmungsdiät bietet die gleichen gesundheitlichen Vorteile des Fastens, ohne tatsächlich zu fasten. Durch den Verzehr von 800 zu 1,100-Kalorien in exakten Mengen und Kombinationen von Lebensmitteln, die für jeden Tag einzeln verpackt und gekennzeichnet wurden, können Sie den menschlichen Körper in einen Fastenzustand versetzen. Durch verschiedene Forschungsstudien entdeckte Dr. Valter Longo, dass unsere Zellen, indem sie den Körper des Essens auf diese Weise berauben, anfangen, unser inneres Gewebe durch einen Prozess, der als Autophagie bekannt ist, zu zerstören und zu regenerieren, wobei geschädigte Zellen getötet, ersetzt oder regeneriert werden. Darüber hinaus kann das Fasten verschiedene Gesundheitsprobleme umkehren, Krebszellen zerstören und die Möglichkeit der Entwicklung einer Alzheimer-Krankheit erheblich reduzieren.
Mit dem Longevity Diet Plan, der im Buch von Dr. Valter Longo vorgestellt wird, werden Sie besser essen, sich besser fühlen und, obwohl er nicht als Gewichtsverlustplan konzipiert ist, vielleicht sogar ein paar Pfunde verlieren. Mit diesem einzigartigen Diätprogramm müssen Sie keine komplexen Ernährungsregeln beachten und schwierige Entscheidungen treffen. Sobald Sie sich mit diesen Änderungen des Lebensstils vertraut gemacht haben, können Sie Ihre allgemeine Gesundheit und Ihr Wohlbefinden verbessern sowie Ihre Langlebigkeit. Das Der Umfang unserer Informationen beschränkt sich auf die Themen Chiropraktik, Wirbelsäulengesundheit und funktionelle Medizin. Um das Thema weiter zu besprechen, wenden Sie sich bitte an Dr. Alex Jimenez oder kontaktieren Sie uns unter 915-850-0900 .
Rückenschmerzen ist eine der häufigsten Ursachen für Behinderungen und versäumte Arbeitstage weltweit. Rückenschmerzen sind der zweithäufigste Grund für Arztbesuche, die nur bei Infektionen der oberen Atemwege unterlegen sind. Ungefähr 80 Prozent der Bevölkerung wird mindestens einmal im Leben Rückenschmerzen haben. Ihre Wirbelsäule ist eine komplexe Struktur, bestehend aus Knochen, Gelenken, Bändern und Muskeln sowie anderen Weichteilen. Verletzungen und / oder erschwerte Bedingungen, wie z Bandscheibenvorfall, kann schließlich zu Rückenschmerzen führen. Sportverletzungen oder Autounfallverletzungen sind oft die häufigste Ursache von Rückenschmerzen, jedoch können manchmal die einfachsten Bewegungen schmerzhafte Folgen haben. Glücklicherweise können alternative Behandlungsmöglichkeiten wie Chiropraktik helfen, Rückenschmerzen durch die Verwendung von Wirbelsäuleneinstellungen und manuellen Manipulationen zu lindern, was letztlich die Schmerzlinderung verbessert.
XYMOGEN s Exklusive professionelle Formeln sind über ausgewählte, lizenzierte Fachkräfte im Gesundheitswesen erhältlich. Der Verkauf und die Diskontierung von XYMOGEN-Formeln im Internet sind strengstens untersagt.
Stolz, Dr. Alexander Jimenez macht XYMOGEN-Formeln nur für Patienten verfügbar, die unter unserer Obhut stehen.
Bitte rufen Sie unser Büro an, um eine ärztliche Konsultation für den sofortigen Zugang zu erhalten.
Wenn Sie ein Patient von sind Injury Medical & Chiropractic Clinickönnen Sie sich über XYMOGEN anrufen 915-850-0900.
Für Ihre Bequemlichkeit und Überprüfung der XYMOGEN Produkte bitte den folgenden Link überprüfen. *XYMOGEN-Katalog-Herunterladen
* Alle oben genannten XYMOGEN-Richtlinien bleiben strikt in Kraft.
Das Find A Practitioner-Tool von IFM ist das größte Überweisungsnetzwerk in der Funktionellen Medizin, das geschaffen wurde, um Patienten dabei zu helfen, Ärzte der Funktionellen Medizin überall auf der Welt zu finden. IFM Certified Practitioner werden aufgrund ihrer umfassenden Ausbildung in Funktioneller Medizin an erster Stelle in den Suchergebnissen aufgeführt